
在过去 50 年,实验室已经能够通过实验演示促成致细胞死亡的过程。早期发现集中于细胞死亡的形态特征以及细胞凋亡和坏死的划分。从那时起,关于导致细胞凋亡的程序性细胞通路有了更多发现。
我们已经通过诸如 胱天蛋白酶 3、bcl-2 和 Fas 等蛋白质进一步理解了凋亡性程序性细胞死亡的基本机制。尽管细胞死亡或许看似简单,但促成细胞死亡的信号转导通路比最初猜想多得多。
受调控坏死性细胞死亡:坏死性凋亡与细胞焦亡
在过去的几年中,在受调控坏死性细胞死亡的研究中取得了令人振奋的进展,其发挥了广泛的生理病理学作用并具有治疗意义。坏死性凋亡和细胞焦亡代表两种共有这些坏死形态特征(包括细胞肿胀、质膜孔形成和膜破裂)的受调控通路。这些过程通过释放众多损伤相关分子模式 (DAMP)(如 HMGB1)和炎性细胞因子(如白细胞介素-1 (IL-1β) 和 IL-18)来触发炎症反应。
坏死性凋亡
坏死性凋亡是在抑制凋亡的条件下激活的细胞防御途径。它需要激活使 MLKL 假激酶磷酸化的 RIPK3 激酶。MLKL 在 Ser358(小鼠中的 Ser345)处磷酸化导致 MLKL 低聚化和成孔复合体形成。MLKL 孔形成阳离子通道,导致进一步的膜破裂和 DAMP 的分泌。RIPK3 激活借助几个 RIP 同型相互作用基序 (RHIM) 结构域相互作用触发,这些基序包括 RIPK1、TRIF 和 ZBP1/DAI,并且导致 RIPK3 在 Ser227(小鼠中的 Thr231/Ser232)处磷酸化。受 TNF 家族成员刺激后,RIPK1 介导了典型的坏死性凋亡信号转导,并导致 RIPK1(包括 Ser166)的自身磷酸化。此通路可以被 RIPK1 的直接抑制剂 necrostatins(Nec-1、Nec-1s)抑制。
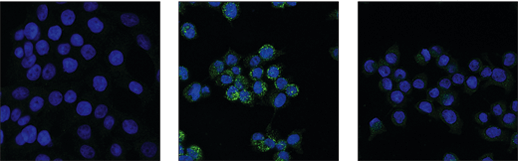 使用 Phospho-RIP3 (Ser227) (D6W2T) Rabbit mAb #93654(绿色)对未经处理(左图)、经 Z-VAD(20 μM,30 分钟)预处理后经 SM-164 (100 nM) 和人肿瘤坏死因子-α (hTNF-α)(20 ng/mL,6 小时;中图)处理、或经 Z-VAD 预处理后经 SM-164 和 hTNF-α 处理并经 λ 磷酸酶后加工(右图)的 HT-29 细胞进行免疫荧光分析。
使用 Phospho-RIP3 (Ser227) (D6W2T) Rabbit mAb #93654(绿色)对未经处理(左图)、经 Z-VAD(20 μM,30 分钟)预处理后经 SM-164 (100 nM) 和人肿瘤坏死因子-α (hTNF-α)(20 ng/mL,6 小时;中图)处理、或经 Z-VAD 预处理后经 SM-164 和 hTNF-α 处理并经 λ 磷酸酶后加工(右图)的 HT-29 细胞进行免疫荧光分析。
或者,RIPK3 可被天然免疫应答(包括Toll 样受体 (TLR) 召集 TRIF)激活或被 DNA 病毒诱导的 ZBP1/DAI 激活而激活。坏死性凋亡在这些情况下激活代表了一种替代性细胞死亡机制,这种机制可以在病原体找到破坏细胞凋亡的途径情况下激活。细胞凋亡通过胱天蛋白酶-8 介导的 RIPK1 和 RIPK3 剪切来抑制坏死性凋亡。坏死性凋亡期间释放的 DAMP 会导致炎症反应,包括激活细胞焦亡。
焦亡
细胞焦亡与坏死性凋亡有一些相似之处,但是虽然认为坏死性凋亡是细胞凋亡受到抑制的继发性细胞死亡反应,但细胞焦亡通常是对传染性生物体的主要反应。细胞焦亡在天然免疫系统的细胞如单核细胞、巨噬细胞和树突状细胞中微生物病原体上表达的病原体相关分子模式 (PAMP) 存在下诱导或被细胞源性 DAMP 诱导。经典的细胞焦亡诱导需要激活胱天蛋白酶-1,这切割并激活炎性细胞因子如 IL-1β 和 IL-18。此外,胱天蛋白酶-1 切割成孔蛋白焦孔素 D (GSDMD)。
探索交互式坏死性细胞死亡通路图
一旦切割,焦孔素 D 的氨基末端片段就低聚化以形成一个类似于但大于 MLKL 的孔,从而允许炎性 DAMP 和细胞因子分泌。在此过程中,胱天蛋白酶 e-1 被各种炎性小复合体激活,这类复合体通常由一个胞质模式识别受体(PRR;一个核苷酸结合结构域和亮氨酸丰富重复序列 [NLR] 或 AIM2 样 [ALR] 家族成员)、一个接头蛋白 (ASC/TMS1) 和原胱天蛋白酶-1 组成。不同炎性体复合体可检测诱发细胞焦亡的不同 PAMP 和 DAMP。由 NLR - NLRP3 触发的最佳表征的通路借助一个两步骤过程发生。第一步骤是一个引发信号,其中激活NF-κB 以诱导众多炎性小体组分表达,这些组分包括 NLRP3、pro-IL-1β 和 pro-IL-18。在第二个激活步骤中,caspase-1 被激活且 gasdermin D 和细胞因子通过蛋白水解被激活。
坏死信号转导通路和疾病
虽然经常在传染病的背景下研究调节坏死的信号转导通路,但事实证明它们具有广泛的病理学意义,包括神经退行性疾病、自身免疫性疾病、癌症、代谢性疾病、心血管疾病、肝脏、胰腺、肾脏和肠道的炎症以及其他慢性炎症性疾病。这些通路的组分可以充当关键生物标记物以及潜在的治疗靶标。
目前,已经实现了靶向 IL-1b 及其受体以治疗关节炎和其他炎性疾病的治疗策略。但是,由于这些方法可导致一般的免疫抑制,因此抑制炎症小体(包括 NLR 和炎性 caspases)的靶向性更强的方法可提供其他广泛适用的治疗方法。还发现了直接靶向于 RIPK1、RIPK3 和 MLKL 的坏死性凋亡的抑制剂。已经对 RIPK1 抑制剂进行了临床研究,该抑制剂可能用于神经退行性疾病和炎性疾病。
总之,这些通路为理解和治疗疾病提供了巨大的希望。
要更多了解参与多种类型细胞死亡的机制、形态和关键蛋白质,请下载《细胞死亡机制的研究者指南》。
坏死性凋亡与细胞焦亡抗体小包装组合
选择参考文献:
- Shan B, Pan H, Najafov A, Yuan J. Necroptosis in development and diseases. Genes Dev. 2018;32(5-6):327-340. doi:10.1101/gad.312561.118
- Frank D, Vince JE. Pyroptosis versus necroptosis: similarities, differences, and crosstalk. Cell Death Differ. 2019;26(1):99-114. doi:10.1038/s41418-018-0212-6
- Shi J, Gao W, Shao F. Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem Sci. 2017;42(4):245-254. doi:10.1016/j.tibs.2016.10.004
- Weinlich R, Oberst A, Beere HM, Green DR. Necroptosis in development, inflammation and disease. Nat Rev Mol Cell Biol. 2017;18(2):127-136. doi:10.1038/nrm.2016.149
- Zakeri Z, Lockshin RA. Cell death: history and future. Adv Exp Med Biol. 2008;615:1-11. doi:10.1007/978-1-4020-6554-5_1


 沪公网安备31011502018823号
沪公网安备31011502018823号