


Toll 样受体(TLR)是跨膜受体,在先天免疫反应中起关键作用。该名称源自与果蝇 Toll 基因的同源性;与在脊椎动物谱系中进化的适应性免疫不同,先天免疫在无脊椎动物分支中存在(并且是保守的)。
这些受体在树突状细胞和巨噬细胞中高度表达,可感知并响应病原体相关的分子模式 (PAMP)(通常在细菌表面上发现),并通过随后的细胞内信号转导通路激活一线防御炎症反应。了解 TLR 信号转导如何被分子模式激活不仅与它们在抵抗感染中的作用有关,而且还与它们在癌症中的作用有关,因为通过损伤相关分子模式 (DAMP) 激活 TLR(宿主细胞中可触发级联反应的生物分子)已被证明可促进肿瘤生长并建立肿瘤对宿主免疫反应的耐受性。
什么是 toll 样受体?
TLR 是在各种组织和许多不同细胞类型中都有表达的单次跨膜受体的超家族。例如,除了它们在免疫细胞中的表达外,在许多上皮组织(可作为抵抗感染的保护层)中也发现了 TLR。相应的,这样的定位使它能对试图穿过上皮的微生物做出快速响应。迄今为止,已经基于常见的 Toll-IL-1 受体(TIR)域确定了 11 个家庭成员。TLR 可以通过其细胞定位进一步分类,如家族成员 TLR1、TLR2、TLR4、TLR5、TLR6 和 TLR11 位于细胞表面,而 TLR3、TLR7、TLR8 和 TLR9 位于内体/溶酶体室。
与通过复杂的体细胞重组建立抗原识别多样性的抗体不同,TLR 通过种系编码的模式识别受体检测病原体。这些受体可识别许多微生物共有的特征(包括细胞表面和细胞壁成分),这些特征可与宿主组织中通常存在的分子区别开来。常见的 PAMP 例子包括脂多糖(LPS)、鞭毛蛋白和肽聚糖(PGN)。包含未甲基化的 CpG 基序的细菌 DNA 也可以通过 TLR 激活诱导先天性免疫反应,从而在数分钟至数小时内产生非常快速的免疫反应时间。
TLR 信号转导和激活
配体结合到 TLR 的胞外域后,通过募集含有 TIR 域的接头蛋白 MyD88,从而在胞质 TIR 域启动信号转导。反过来,MyD88 募集与 IL-1 受体相关的激酶 4 (IRAK-4),它作为通过 IKK 复合物进行下游信号转导的中心枢纽,引起 MAP 激酶(即 JNK 和 p38 MAPK )的激活。不依赖MyD88 的通路激活也可以通过 TRIF 和 TRAF3 发生,从而导致 IKKε/TBK1 募集。最终,通过这些通路的信号转导会汇集至转录因子,包括 IRF7、 IRF3、AP-1 和 NF-kB,导致与免疫反应答细胞因子产生和吞噬作用相关的免疫细胞的存活和增殖相关的基因表达上调。

癌症中的 TLR
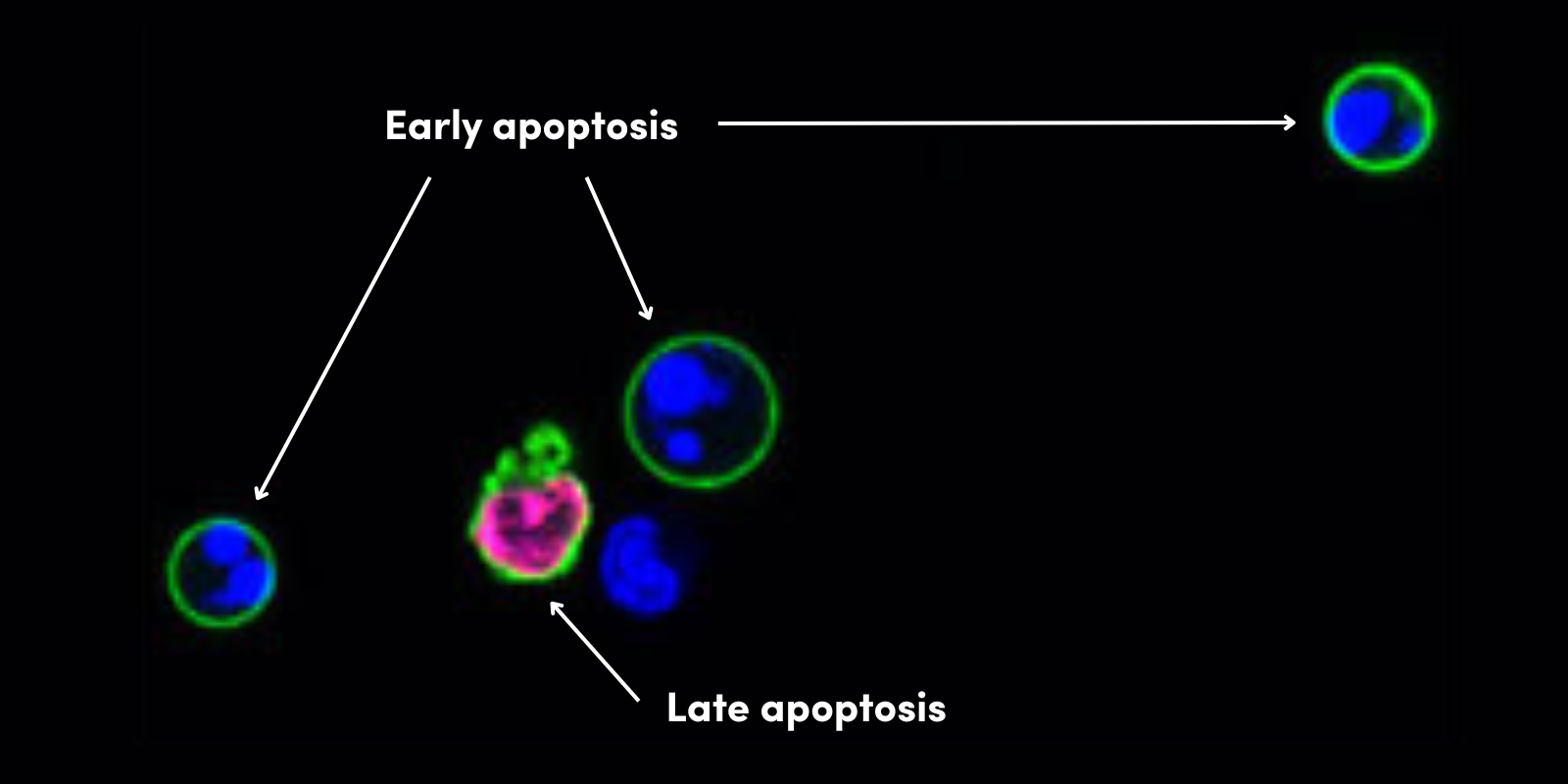
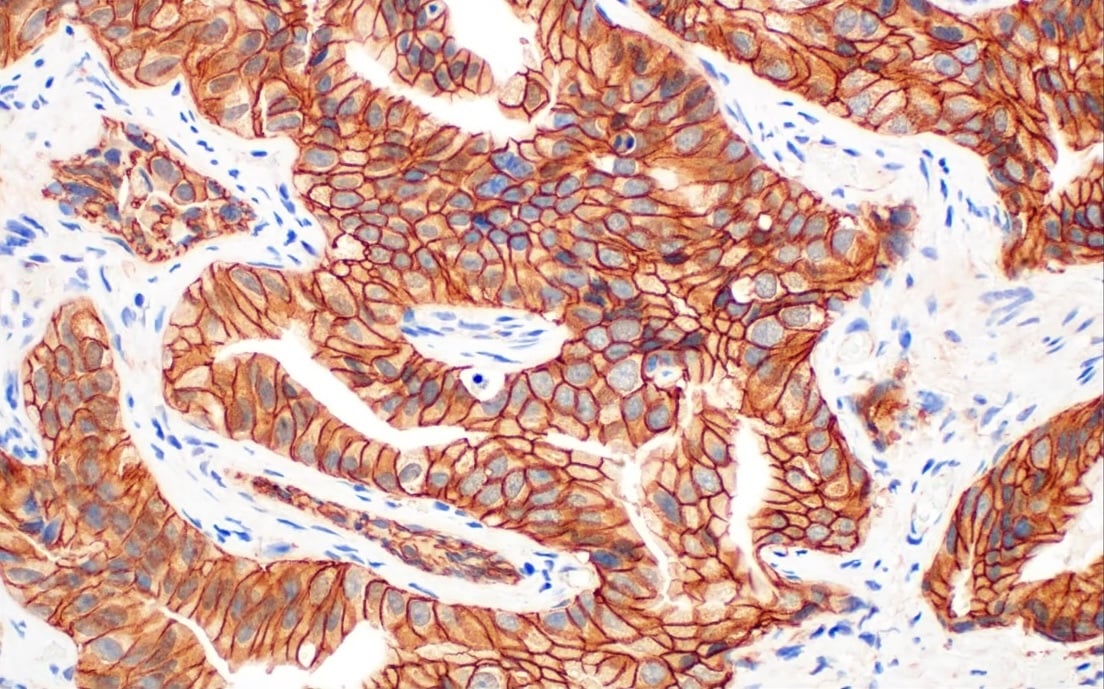
最近,已经报告了在多种癌症(包括脑、肝、前列腺和卵巢肿瘤)中的 TLR 表达。有证据表明,癌细胞中 DAMP 的存在会刺激肿瘤微环境中免疫细胞上的 TLR 激活,从而导致慢性炎症。这些变化与肿瘤进程的改变、凋亡的抑制以及肿瘤对免疫反应的抵抗有关。因此,TLR 激动剂作为抗癌药物具有良好的治疗效果,能够阻止肿瘤的生长和发展。


 沪公网安备31011502018823号
沪公网安备31011502018823号