


线粒体健康是一个复杂且受到精细调节的过程,涉及确定线粒体大小、生物发生和降解的通路。线粒体大小是一个动态过程,涉及融合(扩大)和裂变(收缩)。消除功能失调的线粒体,通常与裂变有关,通过称为线粒体自噬的选择性自噬通路发生。自噬是清除自噬体捕获并被溶酶体降解的细胞质内容物的分解代谢过程。虽然自噬可能是非选择性的,但它也可被用来特异性指向各种细胞器、病原体甚至蛋白质。
线粒体自噬是选择性自噬中的研究最透彻的实例,涉及多个定位于线粒体的货物受体,这些受体能够响应诸如线粒体去极化、氧化应激和缺氧等信号。含有 LC3 相互作用区 (LIR) 的货物受体与 LC3 家族成员相互作用,将目标连接到自噬体,随后被溶酶体降解。
PINK1/Parkin 通路
研究最透彻的线粒体自噬通路涉及在线粒体去极化后激活的 PINK1/Parkin。PINK1 是一种线粒体丝氨酸/苏氨酸激酶,在通过线粒体内膜转运后通常保持在低水平,在那里它被肽酶 PARL 切割并被蛋白酶体降解。抑制线粒体膜电位以防止 PINK1 易位导致线粒体外膜稳定、泛素 Ser65 磷酸化以及 E3 连接酶 Parkin 的募集和磷酸化。
Parkin 的激活会导致多种线粒体蛋白上泛素化链的积累,这些蛋白向泛素结合货物受体发出信号,包括 SQSTM1/p62、Optineurin 和 NDP52。这一过程中同样重要的是货物受体被 TBK1 磷酸化,TBK1 是一种参与先天免疫的激酶,被招募到线粒体并增强线粒体自噬。
除 PINK1 外,目前已鉴定出几种线粒体货物受体,包括 BNIP3、BNIP3L/Nix、FUNDC1、BCL2L13 和 FKBP8。缺氧诱导的线粒体自噬涉及 BNIP3、BNIP3L/Nix 和 FUNDC1。BNIP3 和 BNIP3L/Nix 均由 HIF1α 转录诱导,并含有自噬体招募必需的 LIR 结构域。BNIP3L/Nix 在网织红细胞发育期的线粒体清除中起重要作用。FUNDC1 是另一种对缺氧诱导的线粒体自噬很重要的货物受体,受自噬激酶 ULK1 调控。这些货物受体之间的相互作用正成为研究热点。
相关博客:缺氧及其在代谢中的作用
对疾病和神经退行性病变的影响
过度或不足的线粒体自噬已被描述为导致许多病理状况的因素,包括神经变性、代谢紊乱、肌肉萎缩症、肝脏疾病、心血管疾病和癌症。线粒体自噬缺陷会导致受损线粒体的积累,进而导致活性氧 (ROS) 的积累和细胞死亡。也许最明显的例子是在早发性帕金森病中发现的 PINK1 和 PARK2(编码 Parkin)的基因突变。这些遗传事件表明疾病和线粒体自噬丧失和功能失调的线粒体积累有关。
类似地,OPTN(编码视神经素)、SQSTM1 和 TBK1 基因突变也被发现与肌萎缩侧索硬化 (ALS) 等神经退行性疾病相关。尽管多数病例中线粒体自噬的直接作用尚未明确,但因线粒体自噬不足导致的功能失调线粒体的异常积累确是各类细胞死亡相关疾病的共同特征。
研究线粒体自噬和推进生物标记物发现的检测工具
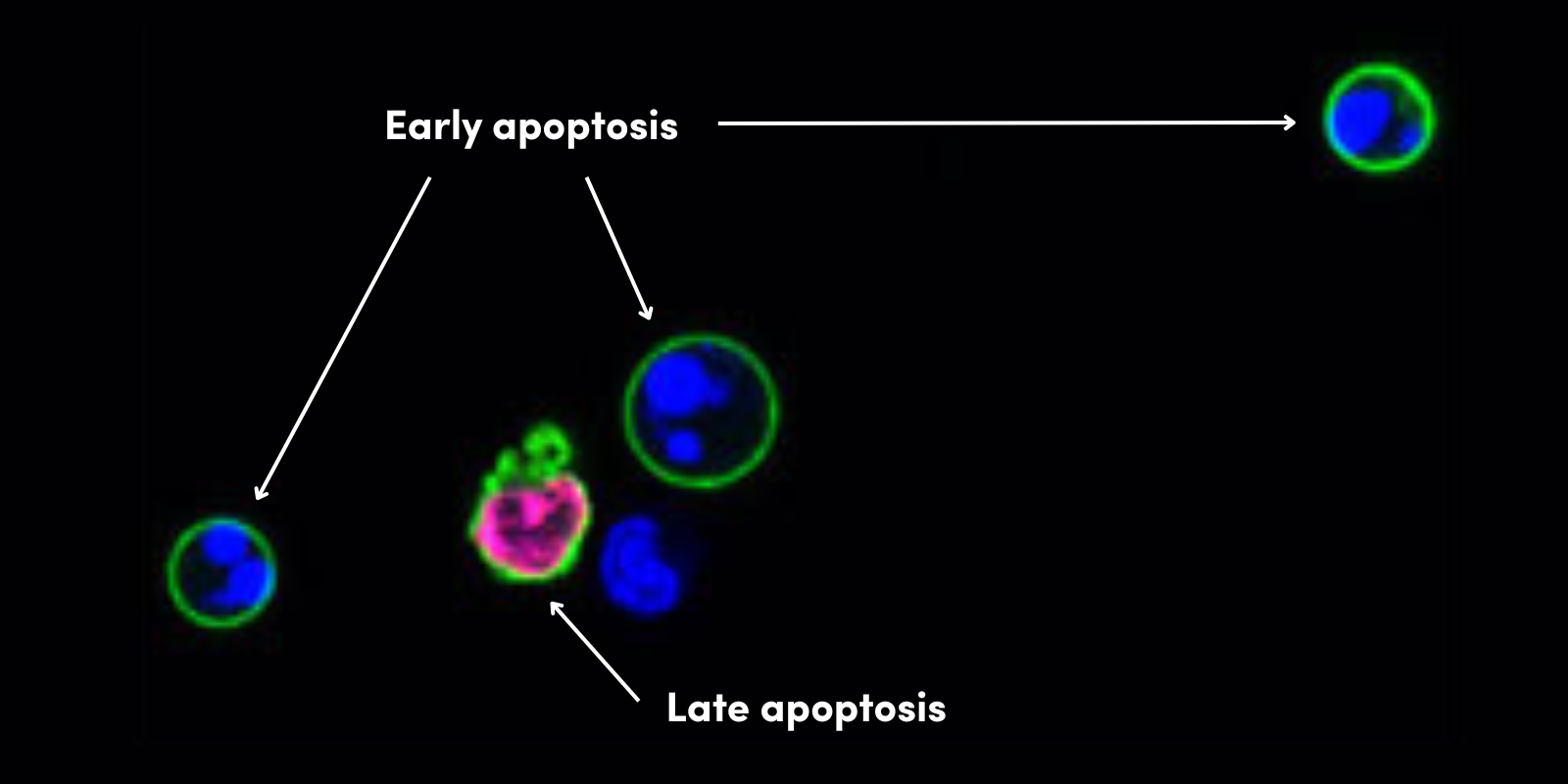
为深入解析线粒体自噬在健康与疾病中的作用,研究人员已开发多种工具来研究这个过程。pH 敏感性线粒体荧光探针 Mt-Keima 与 Mito-QC 虽能用于活体模型中线粒体自噬监测,但对病理样本的适用性有限。此外,通过免疫荧光染色可观察线粒体与自噬体上表达蛋白的共定位情况,但此类方法通常难以实现定量分析。
调控下的线粒体自噬通路的发现为发现有价值的生物标志物和监测线粒体自噬的方法铺平了道路。特异性抗体可用于监测磷酸化 Optineurin 的变化以及 PINK1 及其底物(如泛素)的变化。为检测 磷酸化泛素 (Ser65) 而开发的 ELISA 测定法已被证明具有敏感性和特异性,并且能够测量神经退行性疾病(包括帕金森病和阿尔茨海默病)中发现的线粒体自噬变化。这些测定甚至可能灵敏到可以观察血浆样品中的线粒体自噬差异。
Cell Signaling Technology 开发了经过充分验证的抗体和试剂盒,旨在满足线粒体自噬研究人员的需求并推动治疗发展。


 沪公网安备31011502018823号
沪公网安备31011502018823号