


坏死细胞死亡可能使人联想起一部老的恐怖电影,但与细胞生物学的大多数过程一样,经过更深入的研究,它变得更加令人着迷。
坏死
坏死曾被定义为非程序性的细胞死亡形式,由显著的化学或物理损伤所引起。可引起这种细胞意外死亡的外力包括极端的物理温度、压力、化学应力或渗透压。细胞破裂是坏死的特征,可导致细胞内含物泄漏到细胞外空间。这会导致被称为损伤相关分子模式(DAMP)的生物分子的释放,这些分子可以被免疫细胞识别并触发炎症反应。
探索交互式坏死性细胞死亡通路图
虽然以前曾经认为坏死是被动和非程序性的,但研究发现了类似于坏死的不依赖 caspase 的过程。细胞外信号(死亡受体-配体结合)或细胞内提示(外来微生物核酸),可能会引起某种形式的程序性坏死(称为坏死性凋亡),它能被 caspase 活性显著抑制。
我将在即将发布的博客中介绍许多其他类型的程序性坏死细胞死亡过程,例如焦亡、铁死亡和 NETosis。坏死性细胞死亡的形式(特别是程序性过程)非常重要,因为它们已牵涉到许多疾病的病理学中,包括炎性自身免疫性疾病、纤维化和神经退行性疾病相关的疾病,例如阿尔茨海默氏病(AD)、多发性硬化症(MS)和肌萎缩性侧索硬化症(ALS)。
坏死性凋亡
坏死性凋亡是一种受调节的坏死形式,是一种防止细胞凋亡受阻而被激活的细胞自我破坏的过程。坏死性凋亡不同于凋亡和其他形式的程序性细胞坏死,因为它不依赖于 caspase 活性。相反,它需要 MLKL 的 RIPK3 依赖性磷酸化。这种磷酸化事件使 MLKL 在质膜上产生孔复合体,从而导致 DAMP 的分泌、细胞肿胀和膜破裂。坏死性凋亡期间,可以观察到细胞分解的不同阶段,包括细胞器肿胀、细胞膜破裂,最后是细胞质和细胞核的分解。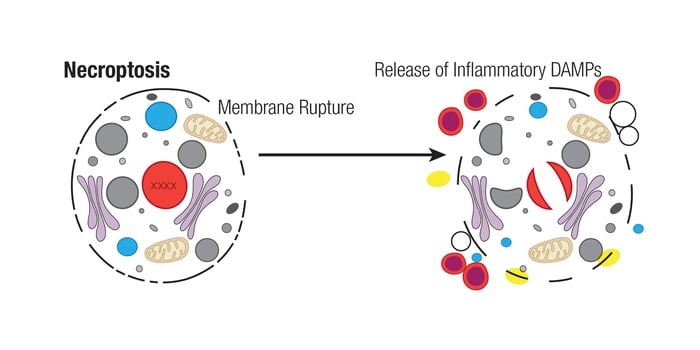
坏死性凋亡本质上是高度免疫原性的,并且可用作针对病毒和其他病原体的宿主防御机制。实验发现其在病毒机制抑制 caspase 活性时发生,从而使细胞自毁以限制病毒复制。在其他病理疾病中也检测到坏死性凋亡,包括缺血性脑损伤、心肌梗死以及与化疗相关的细胞死亡。它还可在炎性疾病、神经退行性和癌症中被激活。
坏死性凋亡通常由细胞外刺激触发。这时候,配体(如 TNF-α)与细胞膜中的死亡受体 (DR) 结合。该 TNF 超家族中的受体包括 TNFR1、Fas/CD95、DR4/TRAIL-R1 和 DR5/TRAIL-R2。一旦被激活,这些受体就与衔接蛋白 TRADD 和 TRAF2 结合,从而导致 RIP 激酶的下游活化。
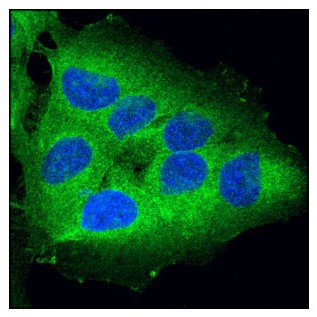
RIP 激酶是调节炎症和细胞死亡的 Ser/Thr 激酶。上图显示使用 RIP (D94C12) XP® Rabbit mAb #3493 (绿色)对 OVCAR8 细胞进行共聚焦 IF 分析。
典型地,在与 RIPK1 形成复合物后,RIPK3 在 Ser227(小鼠中是 Thr231/Ser232)位点通过磷酸化被激活,该复合物本身也在 Ser166、Ser161 和 Ser14/15 位点被磷酸化。这种复合物称为坏死复合物,然后在 Ser358 位点被 MLKL 磷酸化。然后,活化的 MLKL 能够低聚并形成易位至质膜的孔复合物。这些孔复合体在那里与磷脂酰肌醇相互作用,并引起膜通透性和细胞破坏。MLKL 诱导的质膜通透导致 Ca2+ 或 Na + 离子内流并直接形成孔道,释放 DAMP ,例如线粒体 DNA (mtDNA)、HMGB1、白介素 (IL)-33、IL-1α 和 ATP。细胞外空间中炎性 DAMP 的存在是将免疫细胞募集到受损或感染组织的信号。
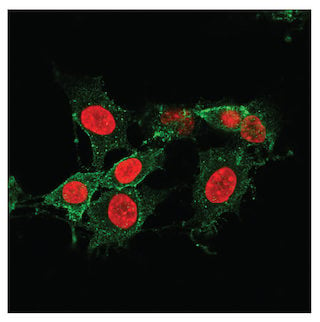 MLKL 的磷酸化可导致孔形成,也是坏死细胞的标记物。上图显示使用 Phospho-MLKL (Ser345) (D6E3G) Rabbit mAb #37333 (绿色)对经过 Z-VAD(20 μM,30 分钟)预处理后再经过 SM-164 (100 nM) 和 Mouse Tumor Necrosis Factor-α (mTNF-α) #5178(20 ng/mL,2.5 小时)处理的 L-929 细胞进行共聚焦免疫荧光分析。
MLKL 的磷酸化可导致孔形成,也是坏死细胞的标记物。上图显示使用 Phospho-MLKL (Ser345) (D6E3G) Rabbit mAb #37333 (绿色)对经过 Z-VAD(20 μM,30 分钟)预处理后再经过 SM-164 (100 nM) 和 Mouse Tumor Necrosis Factor-α (mTNF-α) #5178(20 ng/mL,2.5 小时)处理的 L-929 细胞进行共聚焦免疫荧光分析。
重要的是要注意,RIPK1 是高度调节的,并在启动 NF-kB 信号转导和存活、凋亡或坏死性凋亡的复合物中发挥作用。因此,分析关键蛋白 RIPK1、RIPK3 和 MLKL 的磷酸化状态是识别坏死性凋亡的重要步骤。
坏死性凋亡通路的其他触发因素包括病原体识别受体 (PRR) 的病原体激活,其中包括 Toll 样受体 (TLR),后者激活随后相互作用、不依赖于 RIPK1 而激活 RIPK3 的衔接子 TRIF 和 ZBP-1。
坏死性凋亡的过程可以以各种方式被抑制。由于其常依赖于 RIPK3 和坏死复合物的活性,caspase-8 可通过 RIPK1 和 RIPK3 的裂解抑制坏死。凋亡通路的激活也抑制了坏死。相反,蛋白 FLIP(caspase-8 的无催化活性的同系物)取代了凋亡复合物中的 caspase-8,它可以阻止 RIPK1 的裂解并驱动坏死。Necrostatin-1 (Nec-1) 是一种已被报告可抑制 RIP1K 活性的小分子,已在存在 necrostatin-1 的情况下观察到坏死性凋亡受到抑制。
当前研究提示,坏死性凋亡在癌症以及几种神经退行性疾病中发挥重要作用。坏死性凋亡被认为参与肿瘤转移;因此,抑制坏死性凋亡通路能够限制肿瘤生长。此外,据报道在阿尔茨海默病和帕金森病中,Necrostatin-1 处理能够提高细胞活力。综上所述,研究坏死性凋亡及其他细胞死亡通路的机制可为各种疾病的新型治疗方式开发提供具有治疗价值的见解。
其他资源
要更多了解参与多种类型细胞死亡的机制、形态和关键蛋白质,请下载下面的指南:

阅读细胞死亡机制系列中的其他博客文章:
更多了解 CST® TUNEL 试剂盒,该试剂盒稳健检测发生凋亡的细胞和其他形式的程序性细胞死亡。
参考文献
-
Galluzzi L, Vitale I, Aaronson SA, et al. Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018;25(3):486-541. doi:10.1038/s41418-017-0012-4
- Frank D, Vince JE. Pyroptosis versus necroptosis: similarities, differences, and crosstalk. Cell Death Differ. 2019;26(1):99-114. doi:10.1038/s41418-018-0212-61.
- Escobar ML, Echeverría OM, Vázquez-Nin GH. Necrosis as Programmed Cell Death. In: Cell Death - Autophagy, Apoptosis and Necrosis. InTech.


 沪公网安备31011502018823号
沪公网安备31011502018823号