


病毒感染期间细胞死亡的调节是病毒与宿主存活平衡的重要因素。在本系列中,我们研究了通过细胞对病毒的应答以及对病毒编码蛋白的应答来调节的通路。冠状病毒 SARS-CoV 和 SARS-CoV-2 感染引起严重的肺损伤、肺泡和支气管上皮细胞的广泛受损,并产生肺外损害。

通过病毒感染导致的细胞死亡
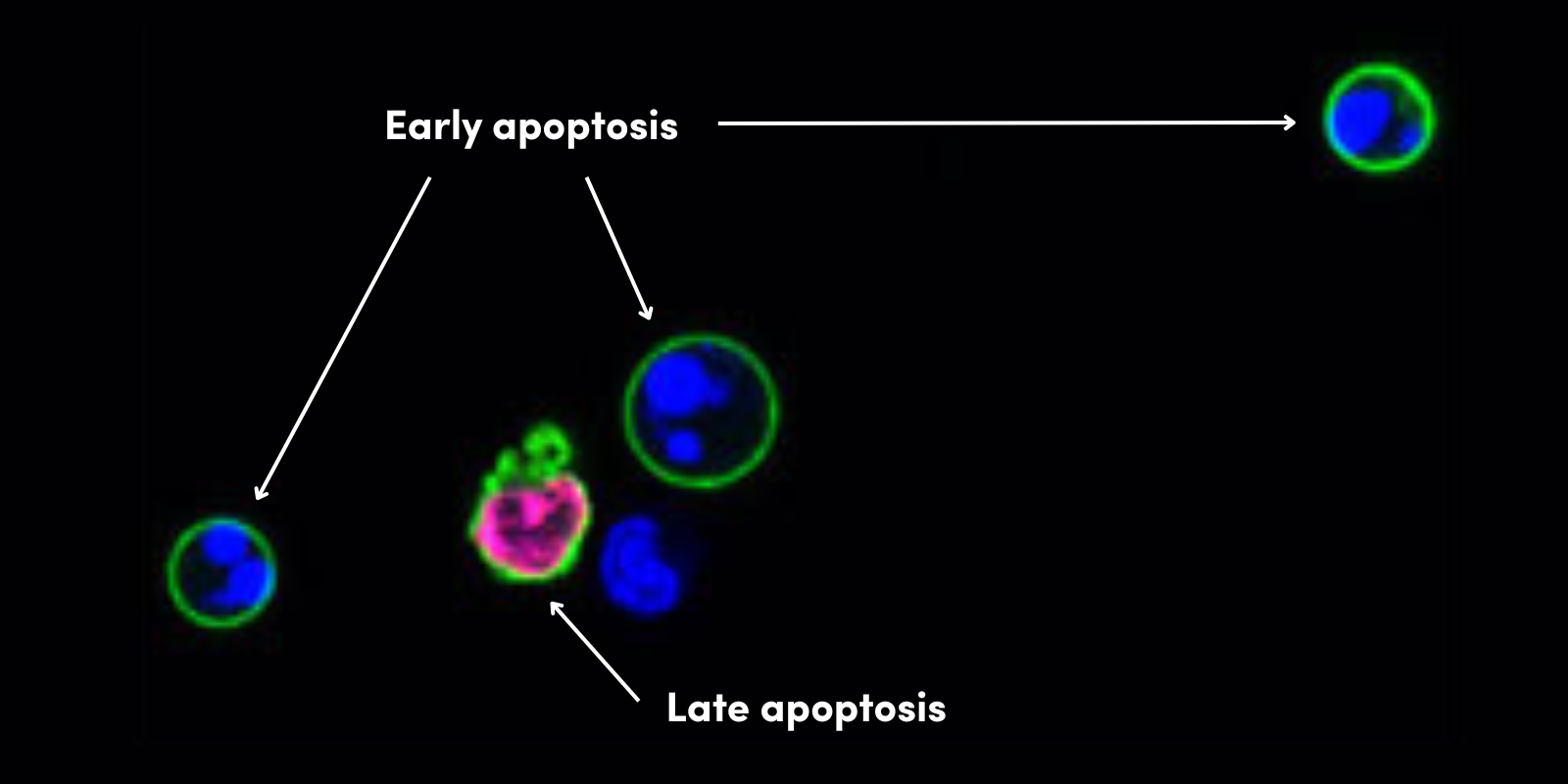
作为对病毒感染的应答,细胞死亡通过炎症细胞因子风暴在受感染细胞和邻近细胞中发生。程序性细胞死亡表现在形态不同的凋亡和坏死过程中,具有高度的信号串扰。
要了解更多关于内质网 (ER) 应激和自噬在病毒感染(特别是 SARS-CoV-2)中的作用,请查看这篇文章。
凋亡可以由几种冠状病毒编码的蛋白质诱导,也可以通过未折叠的蛋白质应答 (UPR) 诱导。这是一个严格控制的过程,研究了几十年,通常与 Caspase-3 的激活有关,后者作为一种酶源产生,通过上游调节性半胱天冬酶的蛋白水解裂解而激活。Caspase-3 能裂解许多参与细胞分解的蛋白质(如 PARP)。
内源性凋亡由 Bcl-2 家族成员控制,可触发线粒体外膜的通透 (MOMP)、细胞色素 c 的释放以及 Caspase-9 和 Caspase-3 的激活。Bcl-2 家族由大量的相关蛋白组成,这些蛋白质包括执行者成员(如 Bax 和 Bak)、抗凋亡成员(Bcl-2、Bcl-xL、Mcl-1、A1/Bfl-1、Bcl-w)和“BH-3 only”蛋白(Bim、Bid、Bad、Bik、Puma、Noxa、Hrk、BMF),它们可高度调节并触发 Bax/Bak 激活。

SARS 冠状病毒已获得直接调控 Bcl-2 家族诱导细胞凋亡的机制。例如,SARS-CoV protein 7a 通过与 Bcl-xL 结合来诱导细胞凋亡。此外,SARS-CoV 和 SARS-CoV-2 包膜蛋白(E)具有一个保守的 BH3 结构域,该结构域可使细胞凋亡,并且对病毒的毒力很重要。
外在通路通过 TNFR 超家族中的死亡受体也可以触发细胞凋亡,从而导致募集 Caspase-8(诱导死亡的信号复合物 (DISC)),并随后激活 Caspase-3。可以通过许多既定测定法监测细胞凋亡,包括末端脱氧核苷酸转移酶 dUTP 缺口末端标记 (TUNEL) 测定法、半胱天冬酶(如 Caspase-3)激活、监测 Bcl -2 家族成员表达、细胞色素 c 释放,以及 Annexin V 染色。

细胞死亡和炎症反应
最近,已经发现坏死细胞死亡可以作为炎症反应的一部分而发生,这很可能导致组织损伤和纤维化。坏死性凋亡和细胞焦亡是代表坏死形态特征的两条通路,这种形态包括细胞肿胀、质膜孔形成和损伤相关分子模式 (DAMP) 的释放,例如 HMGB1 和炎症性细胞因子。
坏死性凋亡是一种细胞防御通路,当凋亡被抑制时会被激活。实际上,许多病毒已经采用了破坏细胞凋亡的机制,从而导致这种裂解反应的激活。它需要将能促进 MLKL (Ser358) 磷酸化 的 RIPK3 激酶激活(在小鼠中 MLKL 的磷酸化位点是 Ser345)。MLKL 的磷酸化导致低聚和孔复合体的形成。RIPK3 激活是通过几个 RIP 同型相互作用基序 (RHIM) 结构域相互作用触发的,包括 RIPK1、TRIF 和 ZBP1,并导致 RIPK3 在 Ser227(小鼠中的 Thr231/Ser232)位点的磷酸化。由 RIPK1 介导的经典性坏死性凋亡信号转导与 RIPK1 在 Ser166 位点上的自磷酸化有关,并可被坏死抑制素抑制。或者,可以通过先天性免疫反应(包括 TRIF 的 Toll 样受体 (TLR) 募集)或通过 DNA 病毒(包括 ZPB1 的激活)来激活 RIPK3。细胞凋亡可通过 Caspase-8 介导的 RIPK1 和 RIPK3 裂解来抑制坏死性凋亡。实际上,在感染了神经侵染性冠状病毒 OC43 的神经元细胞中已经观察到坏死性凋亡。SARS-CoV 辅助蛋白 3a 与 RIPK3 相互作用以帮助推动坏死细胞死亡。

导致坏死性细胞死亡的另一种通路,即脊髓灰质炎,其特征在于 gasdermin 家族成员的孔形成能力。TLR 和 DAMP 激活后,先天免疫系统的细胞中就会引起细胞凋亡。该通路的典型激活涉及 Caspase-1 对 Gasdermin D 的裂解,其中 Gastermin D 的 N 端片段形成膜孔。Caspase-1 还裂解并激活炎症细胞因子(如 IL-1b 和 IL-18),它们是通过 Gasdermin D 孔分泌的。Caspase-1 可以通过称为炎性小体的病原体感应复合体激活。

包含 NLRP3、ASC 和 Caspase-1 的炎症复合体作为涉及微生物病原体、钾外流和溶酶体损伤剂的两步过程的一部分被激活。重要的是,Caspase-3 可以在其他氨基末端位点裂解 Gasdermin D,并使其成孔能力失活。在 SARS 3a 中,如上所述,它激活了坏死性凋亡坏死,但也可以直接或通过 NLRP3 炎性小体激活 caspase-1。研究表明,SARS-CoV 诱导 NLRP3 炎性小体激活和焦亡,并促进血清 IL-1b 水平的升高。
对感染的反应可以包括激活凋亡、坏死的通路,以及在细胞毒力和对宿主组织的损伤中起关键作用的热解。这些通路的串扰程度很高,但是出现了新的摄政者来逗弄这些途径,应该会更好地理解。调节这些反应的新兴疗法的作用是引起人们极大关注的领域。
了解更多:
-
阅读本博客系列的第二部分:自噬和 ER 应激在病毒感染中的作用
-
查看我们关于细胞死亡机制的博客系列:
参考文献
-
Fung TS, Liu DX. Coronavirus infection, ER stress, apoptosis and innate immunity。Front Microbiol. 2014 Jun 17;5:296. doi: 10.3389/fmicb.2014.00296. PMID: 24987391; PMCID: PMC4060729.
-
Tan YX, Tan TH, Lee MJ, Tham PY, Gunalan V, Druce J, Birch C, Catton M, Fu NY, Yu VC, Tan YJ. Induction of apoptosis by the severe acute respiratory syndrome coronavirus 7a protein is dependent on its interaction with the Bcl-XL protein. J Virol. 2007 Jun;81(12):6346-55. doi: 10.1128/JVI.00090-07. Epub 2007 Apr 11. PMID: 17428862; PMCID: PMC1900074.
-
Yue Y, Nabar NR, Shi CS, Kamenyeva O, Xiao X, Hwang IY, Wang M, Kehrl JH. SARS-Coronavirus Open Reading Frame-3a drives multimodal necrotic cell death. Cell Death Dis. 2018 Sep 5;9(9):904. doi: 10.1038/s41419-018-0917-y. PMID: 30185776; PMCID: PMC6125346.
-
Siu KL, Yuen KS, Castaño-Rodriguez C, Ye ZW, Yeung ML, Fung SY, Yuan S, Chan CP, Yuen KY, Enjuanes L, Jin DY. Severe acute respiratory syndrome coronavirus ORF3a protein activates the NLRP3 inflammasome by promoting TRAF3-dependent ubiquitination of ASC. FASEB J. 2019 Aug;33(8):8865-8877. doi: 10.1096/fj.201802418R. Epub 2019 Apr 29. PMID: 31034780; PMCID: PMC6662968.
-
Meessen-Pinard M, Le Coupanec A, Desforges M, Talbot PJ. Pivotal Role of Receptor-Interacting Protein Kinase 1 and Mixed Lineage Kinase Domain-Like in Neuronal Cell Death Induced by the Human Neuroinvasive Coronavirus OC43. J Virol. 2016 Dec 16;91(1):e01513-16. doi: 10.1128/JVI.01513-16. PMID: 27795420; PMCID: PMC5165216.
-
Miller K, McGrath ME, Hu Z, Ariannejad S, Weston S, Frieman M, Jackson WT. Coronavirus interactions with the cellular autophagy machinery. Autophagy. 2020 Sep 23:1-9. doi: 10.1080/15548627.2020.1817280. Epub ahead of print. PMID: 32964796.


 沪公网安备31011502018823号
沪公网安备31011502018823号