


蛋白质腺苷二磷酸或 ADP-核糖基化是一种有趣的翻译后修饰 (PTM),广泛存在于所有生命界中。1 自 Pierre Chambon 60 多年前首次发现该修饰以来,2 科学家们已经表征了 ADP-核糖基化在诸如 DNA 损伤修复和稳态3、细胞骨架组装4、先天免疫5 和 rRNA 生物合成等过程中的相关性。6 由于 ADP-核糖基化在关键细胞过程中发挥重要作用,它已成为一种有前景的治疗干预靶点,并受到广泛关注,尤其是在癌症治疗策略中的应用。
我们最近发布了一种用于检测 ADP-核糖基化修饰的兔单克隆抗体。该产品可按以下几种样式提供:
- Poly/Mono-ADP Ribose (D9P7Z) Rabbit mAb #89190
- Poly/Mono-ADP Ribose (D9P7Z) Rabbit mAb (BSA and Azide Free) #29889
- Poly/Mono-ADP Ribose (D9P7Z) Rabbit mAb (Alexa Fluor® 488 Conjugate) #88873
*一直在增加更多抗体制剂!查看 CST 产品目录,获取此 ADP-核糖抗体克隆的最新产品列表。
与我们的 PTMScan® 产品类似,该克隆可以独立于周围的氨基酸环境识别 ADP 核糖基化 PTM,并且还与物种无关。因此,该克隆可以检测出这种独特的 PTM,无论它出现在谷氨酸、天冬氨酸、丝氨酸、精氨酸、赖氨酸、半胱氨酸还是其他残基上。这种多功能性使其用于研究任何给定生物样品中所有的 ADP-核糖基化情形。
此外,当抗体克隆以单个 ADP-核糖 (ADPr) 单元(称为单 ADP-核糖基化(MAR 或 MARylation))或 ADP-核糖单元链(称为多 ADP-核糖基化(PAR 或 PARylation))的形式附着于蛋白质时,它可以检测到 ADP-核糖基化。
抗原识别基础如此广泛下,如何评估我们抗体克隆的 ADP-核糖基化特异性?本博客介绍了我们在验证测试中进行的实验,包括诱导和抑制 MAR 和 PARylation,您可能会发现这些方法对您自己的研究有帮助。
验证 CST 单聚/多聚 ADP 核糖基化抗体
作为科学家,我们深知拥有一种可以信赖的抗体对于推动研究进展至关重要。开发一种高度特异性的 ADP-核糖基化抗体需要蛋白质组学部门众多研究人员的广泛科学专业知识和共同合作。与所有 CST 抗体验证活动一样,利用多项实验确认抗体特异性,这包括以下三种策略:
- 诱导 MAR/PAR 的二元模型
- 阻断 MAR/PAR 诱导作用的二元模型
- 移除 MAR/PAR 的酶促模型
单独使用这些策略中的任何一种都可以测试抗体特异性,但综合而言,诸如这样的验证活动使我们能够保证 CST 抗体按预期发挥作用。您可以阅读有关以下三个实验的更多信息,其方法可以帮助您了解自己的 MARylation 和 PARylation 实验。
MARylation 和 PARylation 的实验诱导和抑制
为诱导 PAR,我们用过氧化氢 (H2O2) 处理样品。过氧化氢通过 MAPK8(也称为 JNK1)易位到细胞核中来诱导 PAR,在细胞核中,MAPK8 磷酸化并激活生成 PAR 的酶 PARP1。7
相反,他拉唑帕利 处理抑制了 PAR。这种化学物质既充当 PARP 催化抑制剂,又充当 PARP“陷阱”——它将 PAR 生成酶 PARP1 固定至双链 DNA 断口,从而防止 H2O2 介导的 PAR 诱导。3
图 1 中的蛋白质印迹检测显示了诱导 MAR/PAR 和阻断 MAR/PAR 诱导的实验。

图 1.未处理 (-) 或经 H2O2 或他拉唑帕利处理 (+) 的 Hela 细胞的蛋白质印迹分析。
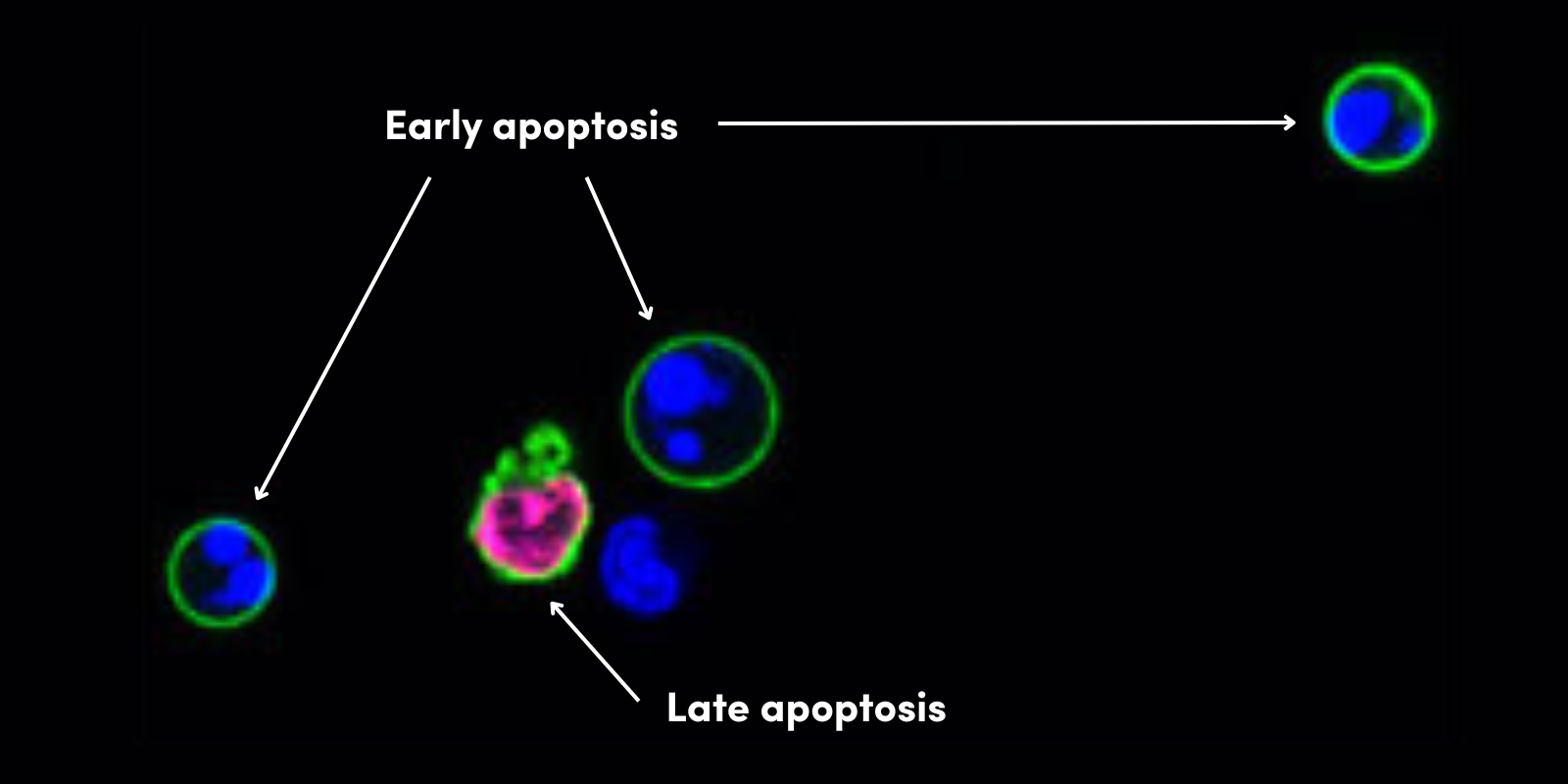
在另一项实验中,聚 ADP 核糖糖基水解酶 (PARG) 用来移除 ADP 核糖基化。PARG 将 PAR 修剪成 MAR,并降低 H2O2 处理的细胞中的 PAR 信号(图 2)。

图 2. 过氧化氢诱导 HeLa 细胞中的 PAR 化信号(左图)并用重组人 PARG 移除 PAR 化信号(右图),其中 IF 检测使用了 Poly/Mono-ADP Ribose (D9P7Z) Rabbit mAb #89190(绿色)、β-Actin (8H10D10) Mouse mAb #3700(红色)和 DAPI #4083(蓝色)。
使用这些实验模型,我们验证了这种新的 PTM 抗体克隆在蛋白质印迹 (WB) 应用和免疫荧光 (IF) 应用中的用途。
治疗潜力:单聚 ADP 核糖基化与多聚 ADP 核糖基化中的 PARP 酶
在诸多胞核过程中,经常强调对 PAR 的调节,因此认为 MAR 在功能上反映 PAR 将是合乎情理的。然而,事实并非如此,某种混淆可能归因于形成这种有趣 PTM 的 PARP 酶家族共享的名称。
聚 ADP 核糖聚合酶 (PARP) 家族的蛋白质共享一个将 ADP 核糖基从 NAD+ 转移到靶标蛋白的催化结构域。存在17 种已知的人 PARP 酶 “撰写器”,它们可以如下诱导 PAR 或 MAR:
- PARP 1、PARP 2、PARP 4、PARP 5A 和 PARP 5B 添加 ADP 核糖的长链 (PAR化)。
- PARP 3、PARP 4、PARP 6、PARP 7、PARP 8、PARP 10、PARP 11、PARP 12、PARP 14、PARP 15 和 PARP 16 参与添加单个 ADP 核糖单元 (MAR 化)。
- PARP4 能够既使底物 MAR 化,又使其 PAR 化。
一个其中 MAR化 与 PAR化不同的情形是在细胞骨架调节中。这里,PARP 7 介导的 α-微管蛋白的 MAR 化导致胞浆中微管解聚。4
探索 CST 产品目录中的相关 PARP 抗体产品。
为了区分 MARylation 和 PARylation,可以采用使用 PARG 共同处理的检测工作流程实验步骤,如图 2 所示。这种处理去除了 PAR 但不去除 MAR,可以与 CST ADP-核糖基化抗体克隆一起在 WB 或 IF 中使用,以进一步研究 MAR 与 PAR 的独特功能。
癌症治疗中的 PARP 抑制剂
在过去十年,PARP 家族已经引起在抗癌战斗中寻找新靶标的肿瘤学家注意。
由于成功的 DNA 修复需要功能性 PAR 化,因此可以在联用 PARP 抑制剂诱导双链断口的癌症治疗(如铂基疗法)中利用这种要求。这使 DNA 在遭停滞的复制分叉处未修复,并诱导基因组不稳定性。
多项临床试验已展示 PARP 抑制剂在胰腺癌、小细胞肺癌以及卵巢癌和乳腺癌中的效力,无论 BRCA1 状态如何。9 截至 2024 年,美国 FDA 已批准四种 PARP 抑制剂(奥拉帕利、卢卡帕利、尼拉帕利 和 他拉唑帕利)用于治疗卵巢癌、输卵管和原发性腹膜癌以及 HER-2 阴性乳腺癌。
作为具有深远临床意义的生物学核心 PTM 之一,看看 ADP 核糖基化研究引领我们走向何方将令人期待。
相关产品和进一步阅读
探索 CST 的相关 PARP 抗体产品,以助力您的 MAR 化或 PAR 化研究:
- PARP (46D11) Rabbit mAb #9532(经验证用于 WB、IP 和 eClip)
-
PARP Antibody #9542(经验证用于 WB)
-
PARP (46D11) Rabbit mAb (BSA and Azide Free) #17245(经验证用于 WB)
-
PARP (46D11) Rabbit mAb (Sepharose® Bead Conjugate) #6704(经验证用于 IP)
请阅读以下博客,更多了解 CST 基序技术:蛋白质组学分析的革命:20 年后的 PTMScan
--
CST 蛋白质组学部门的首席科学家 Barry M. Zee 博士和 Jeff Silva 博士以及科学内容营销经理 Alexandra Foley 参与撰写了这篇博文。
参考文献
- Perina D, Mikoč A, Ahel J, Ćetković H, Žaja R, Ahel I. Distribution of protein poly(ADP-ribosyl)ation systems across all domains of life. DNA Repair (Amst). 2014;23:4-16. doi:10.1016/j.dnarep.2014.05.003
- Chambon P, Weill JD, Mandel P. Nicotinamide mononucleotide activation of a new DNA-dependent polyadenylic acid synthesizing nuclear enzyme. Biochem Biophys Res Commun. 1963;11:39-43. doi:10.1016/0006-291x(63)90024-x
- Rouleau-Turcotte É, Pascal JM. ADP-ribose contributions to genome stability and PARP enzyme trapping on sites of DNA damage; paradigm shifts for a coming-of-age modification. J Biol Chem. 2023;299(12):105397. doi:10.1016/j.jbc.2023.105397
- Palavalli Parsons LH, Challa S, Gibson BA 等人 Identification of PARP-7 substrates reveals a role for MARylation in microtubule control in ovarian cancer cells. Elife. 2021;10:e60481. Published 2021 Jan 21. doi:10.7554/eLife.60481
- Fehr AR, Singh SA, Kerr CM, Mukai S, Higashi H, Aikawa M. The impact of PARPs and ADP-ribosylation on inflammation and host-pathogen interactions. Genes Dev. 2020;34(5-6):341-359. doi:10.1101/gad.334425.119
- Kim DS, Challa S, Jones A, Kraus WL. PARPs and ADP-ribosylation in RNA biology: from RNA expression and processing to protein translation and proteostasis. Genes Dev. 2020;34(5-6):302-320. doi:10.1101/gad.334433.119
- Zhang S, Lin Y, Kim YS, Hande MP, Liu ZG, Shen HM. c-Jun N-terminal kinase mediates hydrogen peroxide-induced cell death via sustained poly(ADP-ribose) polymerase-1 activation. Cell Death Differ. 2007;14(5):1001-1010. doi:10.1038/sj.cdd.4402088
- Slade D, Dunstan MS, Barkauskaite E 等人 The structure and catalytic mechanism of a poly(ADP-ribose) glycohydrolase. Nature. 2011;477(7366):616-620. Published 2011 Sep 4. doi:10.1038/nature10404
- Slade D. PARP and PARG inhibitors in cancer treatment. Genes Dev. 2020;34(5-6):360-394. doi:10.1101/gad.334516.11


 沪公网安备31011502018823号
沪公网安备31011502018823号